
 כניסה
כניסה  עקבו אחרינו בפייסבוק
עקבו אחרינו בפייסבוק הבדלים בין גרסאות בדף "חומצות מרה - Bile acids"
| שורה 129: | שורה 129: | ||
</gallery> | </gallery> | ||
| − | ===המתווה והפיקוח של הפרשת המרה=== | + | ===המתווה והפיקוח של הפרשת המרה=== |
| − | + | הפרשת המרה נמוכה ביותר בעת צום, כאשר רוב נוזל המרה מוסט לכיס המרה לצורך ריכוזו. כאשר בליל המזון המעוכל בחלקו בצאתו מהקיבה לתריסריון הידוע כ-chyme נכנס למעי הדק, חומצה ושומני מזון וחלבונים המעוכלים באופן חלקי, מעודדים הפרשה של 2 הורמונים, [[cholecystokinin]] או CCK ו-secretin. הורמוני מעי אלה משפיעים באופן משמעותי על הפרשת הורמוני הלבלב, ובנוסף יש להם תפקיד בהפרשת ושפיעת המרה לתוך המעי: הגירוי המשמעותי ביותר להפרשת cholecystokinin היא נוכחות שומנים בתריסריון, ואילו הגירוי המשפיע על הפרשת secretin היא דרגת החומציות בתריסריון. השפעת secretin על מערכת המרה היא בגירוי של תאים בצינורות המרה להפריש ביקרבונאט ומים, מה שמרחיב את נפח המרה ומגדיל את קצב שפיעתה אל המעי. | |
| − | הפרשת המרה נמוכה ביותר בעת צום, כאשר רוב נוזל המרה מוסט לכיס המרה לצורך ריכוזו. כאשר בליל המזון המעוכל בחלקו בצאתו מהקיבה לתריסריון הידוע כ-chyme נכנס למעי הדק, חומצה ושומני מזון וחלבונים המעוכלים באופן חלקי, מעודדים הפרשה של 2 הורמונים, | ||
שתי חומצות המרה השכיחות ביותר בנוזל המרה הן chenodeoxycholic acid (המהווה 45%) ו-cholic acid (המהווה 31%). לפני ש-2 חומצות מרה ראשוניות אלו מופרשות לתוך הלומן הקניקולארי, הן עוברות קוניוגציה דרך הקשר האמידי בקצה הקרבוקסי-טרמינאלי שלהן עם גליצין או עם טאורין. קוניוגציה זו מגבירה את האופי האמפיפאתי שלהן, מה שמאפשר להן להיות מופרשות ביתר קלות ואף להיות פחות טוקסיות. קוניוגאטים של חומצות מרה הם המרכיבים העיקריים של נוזל המרה. | שתי חומצות המרה השכיחות ביותר בנוזל המרה הן chenodeoxycholic acid (המהווה 45%) ו-cholic acid (המהווה 31%). לפני ש-2 חומצות מרה ראשוניות אלו מופרשות לתוך הלומן הקניקולארי, הן עוברות קוניוגציה דרך הקשר האמידי בקצה הקרבוקסי-טרמינאלי שלהן עם גליצין או עם טאורין. קוניוגציה זו מגבירה את האופי האמפיפאתי שלהן, מה שמאפשר להן להיות מופרשות ביתר קלות ואף להיות פחות טוקסיות. קוניוגאטים של חומצות מרה הם המרכיבים העיקריים של נוזל המרה. | ||
גרסה מ־11:54, 5 באפריל 2017
| מדריך בדיקות מעבדה | |
| חומצות מרה | |
|---|---|
| Bile acids | |
| מעבדה | כימיה בדם, גסטרו |
| תחום | הערכת מפגעים בכבד או בכיס המרה |
 | |
| טווח ערכים תקין | בנסיוב לאחר צום של 12 שעות רמת סך חומצות מרה - 0.73-5.63 מיקרומול' לליטר (0.3-2.3 מיקרוגרם למ"ל); בנסיוב שעה אחת לאחר ארוחה - 4.41-7.83 מיקרומול' לליטר (1.8-3.2 מיקרוגרם למ"ל); בנסיוב שעתיים לאחר ארוחה - 6.3-16.7 מיקרומול' לליטר (2.57-6.82 מיקרוגרל למ"ל); בצואה - 5-10 מיליגרם לגר', או 120-225 מיליגרם ליום (294-551 מיקרומול' ליום); בשתן - 100-400 מיקרוגרם ל-24 שעות; בנוזל המרה - 15-30 מיליגרם למ"ל; בדם פורטאלי - 10-20 מיקרוגרם למ"ל.
חומצה כולית: בנסיוב לאחר צום של 12 שעות - 0.12-0.61 מיקרוגרם למ"ל (0.3-1.5 מיקרומול' לליטר); בנוזל המרה - 5-20 מיליגרם למ"ל (12-48 מיקרומול' לליטר); בשתן - 5-25 מיקרוגרם ל-24 שעות (12-61 ננומול' ל-24 שעות); בצואה - 0.5-2.0 מיליגרם לגר' (1.2-4.9 מיקרומול' לגרם); בדם הפורטאלי - 5-10 מיקרוגרם למ"ל (12-24 מילימול' לליטר). חומצה כנודאוקסיכולית (chenodeoxycholic acid): בנסיוב לאחר צום של 12 שעות - 0.16-0.98 מיקרוגם למ"ל (0.4-2.5מיקרומול' לליטר ; בנוזל המרה - 5-15 מיליגרם למ"ל; בשתן - 20-100 מיקרוגרם ל-24 שעות; בצואה - 0.2-1.0 מיליגרם לגר'; בדם הפורטאלי - 5-8 מיקרוגרם למ"ל. רמה ממוצעת של חומצות מרה נוספות בנסיוב בצום (בערכי מיקרוגרם למ"ל):
|
| יוצר הערך | פרופ' בן-עמי סלע |
מטרת הבדיקה
הכבד סופג חומצות מרה ביעילות מהצירקולציה הפורטאלית, כתוצאה מהנוכחות של טרנספורטרים של חומצות מרה על פני ממברנות סינוסוידאליות של הפאטוציטים. היעילות הגבוהה של יכולת מיצוי חומצות מרה על ידי הכבד, היא הסיבה לריכוז נמוך יחסית של חומצות אלה בדם ההיקפי (2-6 מיקרומול' לליטר), בהשוואה לריכוזן בצירקולציה הפורטאלית שיכול להגיע ל-60-80 מיקרומול' לליטר. כל ירידה ביכולת המיצוי של חומצות מרה על ידי הכבד, הנגרמת על ידי הפחתה בשפיעת הדם בכבד, או כתוצאה מנזק הפאטו-צלולרי, תגרום לעלייה ברמת חומצות מרה בנסיוב.
לכן, רמות חומצות מרה בנסיוב או בפלזמה, הם מדדים רגישים לתפקוד הכבד, המשקפות את הסינתזה של חומצות אלה בכבד, את הפרשתן ואת ספיגתן מחדש. מדידת רמת חומצות מרה בנסיוב תסייע לגלות שינויים בתפקוד הכבד עוד לפני הופעת תסמינים מתקדמים יותר של תחלואה כגון צהבת, ותעודד טיפול מוקדם עוד לפני יצירת נזק כבדי בלתי הפיך. מחקרים באדם מראים שבמפגעי כבד שונים מדידת רמת חומצות מרה מספקת מידע שאינו מתקבל ממדדי תפקוד כבד מקובלים כגון AST ו-ALT. בעוד ששני האנזימים האחרונים מופרשים מתאי כבד שניזוקו ולכן הם משקפים פגיעה הפאטו-צלולרית, הרי שרמת חומצות מרה היא מדד לתפקוד כבד, למצבי עימדון (cholastasis) הכבד כתוצאה ממפגע חסימתי, ממפגעי כיס המרה כתוצאה מיצירת אבנים ועוד.
מדידת רמת חומצות מרה בדם רלוונטית לאבחון תרחישים כגון portosystemic shunt וכן hepatic microvascular dysplasia. בכל הצורות של פגיעה בתפקוד הפאטו-ביליארי רמת חומצות המרה הנמדדת בנסיוב לאחר הארוחה (postprandial) תימצא גבוהה מסף הנורמה העליון המקובל. לכן מדידת חומצות מרה 1-2 שעות לאחר ארוחה היא מדד מדויק יחסית למפגע הפאטו-ביליארי. מדידת רמת חומצות מרה בנסיוב לאחר ארוחה, היא בעל רגישות וספציפיות של למעלה מ-98% בגילוי של מגוון מפגעים הפאטו-ביליאריים , זאת למרות שרמת חומצות מרה ב-40% מתוך אותם מטופלים נמצאה תקינה.
אחד התרחישים המובהקים המחייב מדידת רמת חומצות מרה בדם, הוא זה של ICP או intrahepatic cholestasis of pregnacy, המתבטא בעיקר בהופעת גרד (pruritus) חמור בכל הגוף. ריכוז יתר של חומצות מרה במעי הגס גורם לשלשולים כרוניים. תרחיש זה של bile acid malabsorption, יכול להיות מאובחן על ידי מבחן SeHCAT או23-selenium-25-homocholic acid taurine , ולהיות מטופל על ידי bile acid sequestrants.
בסיס פיזיולוגי
העבודה החלוצית על המבנה המולקולארי והפיזיולוגיה של חומצות מרה נעשתה בשנות ה-20 באוניברסיטת מינכן, על ידי Heinrich Otto Wieland, והוא זכה על עבודתו זו בפרס נובל בכימיה בשנת 1927.
חומצות מרה מרכיבות משפחה גדולה של מולקולות המורכבות ממבנה בסיסי סטרואידי עם 4 טבעות, ושרשרת צדדית של 5 או 8 פחמנים המסתיימת בקצהה בחומצה קרבוקסילית, וכן מספר שונה של שיירי הידרוקסיל. ארבעת הטבעות מסומנות משמאל לימין כ-A,B,C,D, כאשר טבעת D אינה משושה אלא מכילה 5 פחמנים. שיירי ההידרוקסיל יכולים להיות בתצורת בטא או אלפא, כאשר כל חומצות המרה מכילות שייר הידרוקסיל בעמדה 3 הנובע ממולקולת האם, כולסטרול. במינים רבים, השלב הראשון ביצירת חומצת מרה, הוא הוספת שייר הידרוקסיל בעמדה 7. בשלב הבא של הפיכת כולסטרול לחומצת מרה, משתנה נקודת הצומת בין 2 הטבעות הסטרואידיות, A ו-B, מה שגורם לכיפוף המולקולה, ובתהליך זה שייר 3-הידרוקסיל מותמר לתצורת α.
חומצות מרה הן חומצות סטרואידיות המכילות 24 פחמנים הנוצרות מכולסטרול בכבד. חומצות מרה מופיעות בעיקר במיץ המרה ביונקים. כל חומצות המרה המופרשות מהכבד עברו קוניוגציה עם חומצת אמינו, גליצין או טאורין, והאחרונות יוצרות קומפלקס עם קטיון, בדרך כלל נתרן, והופכות למלחי מרה. באדם מלחים של taurocholic acid ושל glycocholic acid (שניהם נגזרי cholic acid), מייצגים בערך 80% מכלל מלחי המרה. חומצות מרה, נגזרות glycine ו-taurine של חומצות מרה, וכן שתי הנגזרות deoxycholic acid ו-lithocholic acid המכילות שייר הידרוקסיל בעמדה 7α, כל אלה נמצאות באדם בנוזל המרה במעיים.
תפקידן העיקרי של חומצות מרה הוא לסייע ביצירת מיצלות, המקלות על ההכנה לעיכול של שומנים במזון. חומצות מרה הן מולקולות אמפיפטיות מה שמאפשר להן לתחלב על ידי פעילות דטרגנטית חלקיקי שומן גדולים מהמזון, ולהפכם ל"טיפות" זעירות בגודלן. בכך גדל שטח הפנים של השומן, מה שחושף אותו לפעילות המעכלת של lipase, אנזים שאינו מסוגל לחדור אל תוך חלקיקי השומן.
חמש חומצת מרה מהוות למעלה מ-99% מכלל מאגר חומצות המרה בנוזלי הגוף: הכבד מסנתז 2 חומצות מרה ראשוניות: Cholic acid (המהווה כ-31% מתכולת חומצות המרה), שמשקלה המולקולארי 408.5 והרכבה 3α,7α,12α-trihydroxy-5β-cholanic acid, וכן chenodeoxycholic acid (המהווה 45% מתכולת חומצות המרה) שמשקלה המולקולארי 392.5 והרכבה 3α,7α-dihydroxy-5β-cholanic acid. שתי חומצות אלה עברו התמרה על ידי חיידקי מעי לשתי חומצות מרה שניוניות, deoxycholic acid (המהווה 26-28% מתכולת חומצות המרה) שהרכבה 3α,12α-dihydroxy-5β-cholanic acid ומשקלה המולקולארי 392.5, ו-lithocholic acid (המהווה 1-2% מתכולת חומצות המרה) שהרכבה 3α-hydroxy-5β-cholanic acid, שמשקלה המולקולארי 376.5. חומצת מרה חמישית-ursodeoxycholic acid (המהווה פחות מ-1% מתכולת חומצות המרה), שהיא חומצה שלישונית, נוצרת בכבד מהתמרה של lithocholic acid, שהרכבה 3α,7β-dicholanic acid, ומשקלה המולקולארי 392.5.
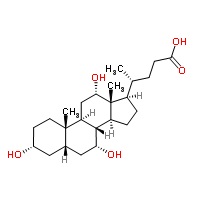
Cholic acid
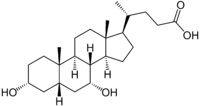
Chenodeoxycholic acid
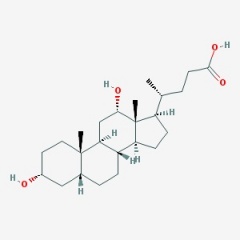
Deoxycholic acid
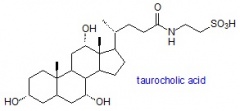
Glycocholic acid






כאמור, באדם הסינתזה של חומצות מרה מתחילה כאשר תאי כבד מסנתזים 2 חומצות מרה ראשוניות, cholic acid ו-chenodeoxycholic acid, על ידי חמצון של כולסטרול במערכת ציטוכרום P-450. מדי יום מסונתזים בערך 600 מיליגרם של מלחי מרה כדי למלא את מקומם של מלחי מרה האובדים בצואה. באדם, הצעד המגביל בהוספת שייר הידרוקסיל לעמדה 7 של גרעין הסטרואיד הוא האנזים cholesterol 7α-hydroxylase, שפעילותו מדוכאת על ידי חומצה כולית, אך מעודדת על ידי כולסטרול.
כאשר 2 חומצות המרה הראשוניות מופרשות למעי, חיידקי-מעי מסירים שיירי OH- מ-cholate ומ-chenodeoxycholate, ליצירה של חומצות המרה השניוניות deoxycholate ו-lithocholate, בהתאמה. כל 4 החומצות הללו יכולות לחזור אל זרם הדם, וממנו לכבד, ולהיות מופרשות מחדש בתהליך הידוע כ-enterohepatic circulation.
החומצה chenodeoxycholic נוצרת על ידי מינים רבים, והיא שימושית ביותר. החיסרון העיקרי של חומצה זו נעוצה ביכולת של חיידקי מעי להסיר את שייר ה-7α hydroxyl באופן שחומצת המרה המתקבלת מכילה רק שייר 3α hydroxyl שידועה כ-lithocholic acid, כשהביטוי litho (אבן) מרמז למסיסות הנמוכה של חומצה זו, ובכך היא טוקסית לתאים. כדי להימנע מבעיות הכרוכות ביצירה של חומצה ליטוכולית, רוב המינים מוסיפים שייר הידרוקסילי שלישי ל-chenodeoxycholic acid. בדרך זו, הרחקת השייר ה-7α הידרוקסילי על ידי חיידקי המעי, מביאה ליצירת חומצת מרה פחות טוקסית אך עדיין פונקציונאלית.
בתהליך האבולוציה של בעלי חוליות, מספר עמדות על פני השלד הסטרואידי נבחרו למיקום השייר ההידרוקסילי השלישי. בתחילה, הועדפה עמדה 16αבעיקר בעופות, ובהמשך עמדה זו פנתה מקומה בסולם ההעדפות לעמדה 12α על ידי מיני בעלי חיים רבים. פרימאטים, כולל האדם, משתמשים בעמדה 12α כמיקום של שייר ההידרוקסיל השלישי. באדם, חומצת המרה הראשונית עם שיר הידרוקסילי בעמדה 12α היא חומצה כולית.
קודם להפרשה של כל אחת מ-4 חומצות המרה הללו, תאי הכבד יכולים לקשור להן אחת משתי חומצות האמינו, glycine או taurine, ליצירה של 8 חומצות מרה מותמרות (conjugated) אפשריות, שהן למעשה מלחי מרה. ה-pKa של כל אחד ממלחי מרה אלה הוא 7.0, כאשר ה-pH של התריסריון הוא בין 3.0-5.0, כך שכאשר חומצות מרה בלתי מותמרות נמצאות בתריסריון, הן תמיד מופיעות בצורתן החומצית [protonated או צורת (HA)], שהיא יחסית צורה בלתי מסיסה במים. תהליך הקוניוגציה של חומצות אלה עם אחת משתי חומצות האמינו שהוזכרו, מוריד את ה-pKa של הקוניוגאט, לתחום של 1.0 עד 4.0. לכן, קוניוגאטים של חומצות מרה הם כמעט תמיד בתצורה המשוללת מימן [deprotonated או (-A)] בתריסריון, שהיא צורה מסיסה בהרבה במים ומסוגלת למלא את התפקיד הפיזיולוגי של חומצות המרה בתחלוב (emulsification) של שומנים. חומצות מרה הן מולקולות פעילות-שטח, דהינו מורידות את מתח הפנים ובכך יכולתן לתחלב שומנים, שלב חיוני ביצירת מיצלות.
דרך אחת בה מסיסות-יתר זו מסייעת לתפקוד של מלחי מרה, היא במניעה של ספיגה-מחדש פסיבית לדם, לאחר שחומצות המרה הופרשו למעי הדק. כתוצאה מכך, הריכוז של חומצות ומלחי מרה במעי הדק יכול להישאר גבוה דיו ליצירת מיצלות ולהמסת שומנים.
לחומצות מרה יש תפקידים נוספים, כולל הרחקת כולסטרול מהגוף, הכוונת הזרימה של המרה כדי להרחיק תוצרי פירוק של מרה מהכבד, תחלוב של ליפידים וויטמינים מסיסי שומן במעי ליצירת מיצלות שיכולות להיות מועברות דרך מערכת הקפילארות הלימפטיות (lacteal system) הסופגת שומנים מהמזון בסיסים (villi) של המעי הדק, וסיוע בהפחתת פלורת החיידקים במעי הדק ובצינורות המרה.
כמות קטנה של חומצות מרה מופרשת דרך הכליות וניתן למצאן בשתן. למרות שהמעיים הם המסלול העיקרי להפרשת חומצות מרה, הפרשת חומצות מרה בשתן משמעותית יותר במפגעים הפאטו-ביליאריים כגון במחלת כבד חסימתית. הכבד עצמו יכול לעודד הפרשת חומצות מרה דרך הכליות במצבים קליניים אחדים, והדבר מושג על ידי תהליך של סןלפציה כאשר הכבד מוסיף שייר סולפאט לחומצת שומן שכבר עברה קוניוגציה, ובכך הופך אותה ליותר קוטבית ומסיסה לצורך פינוי כלייתי.
סינתזה ושימוש בחומצות מרה
חומצות מרה מהוות תוצר סופי של שימוש בכולסטרול בגוף. סינתזה של חומצות מרה היא מסלול עיקרי במטבוליזם של כולסטרול ברוב המינים פרט לאדם. הגוף מייצר מדי יום כ-800 מיליגרם כולסטרול, כאשר מחצית מכמות זו מנוצלת לסינתזה של חומצות מרה. מדי יום מופרשים למעי כ-20-30 גרם חומצות מרה, כאשר כ-90% מכמות זו נספגת מחדש בטרנספורט אקטיבי במעי העקום (ileum), ועוברים מִחזוּר בחזרה לכבד ולכיס המרה. תהליך זה מאפשר קצב נמוך של סינתזה יומית של חומצות מרה, אך יחד עם זאת הפרשה גדולה שלהן למערכת העיכול.
למרות שמספר אנזימים הכרוכים בסינתזה של חומצות מרה פעילים בסוגי תאים רבים, הכבד הוא האיבר היחיד בו מתקיימת הסינתזה המלאה של חומצות מרה. למעשה, סינתזת חומצות מרה מהווה מנגנון עיקרי של הפרשת עודף כולסטרול, אולם הפרשת כולסטרול בצורת מלחי מרה אינה מספיקה לפצות על עודף צריכת כולסטרול בדיאטה. למרות שסינתזת חומצות מרה יוצרת מסלול קטבולי של כולסטרול, חומצות אלה חיוניות גם להמסה של כולסטרול במזון, כמו גם של ליפידים אחרים ומרכיבים חיוניים כוויטמינים מסיסי שומן, כשלב חיוני בהעברת מרכיבי מזון אלה לכבד.
סינתזה של כלל חומצות המרה דורשת פעילות של 17 אנזימים, ומתרחשת בציטופלזמת התאים, ברטיקולום האנדופלזמי (ER), במיטוכונדריה ובפראוקסיזומים. בגלל העובדה שרבים מתוצרי המטבוליזם של חומצות מרה הם טוקסיים לתא, מובנת הבקרה המוקפדת על ייצור חומצות אלה, אכן, מספר מפגעים מולדים במטבוליזם של חומצות מרה, נגרמים מפגמים בגנים שאחראיים לסינתזת חומצות מרה, כאשר מפגעים אלה קשורים לכשל כבדי כבר בגיל הרך, וחלקם מתפתחים לנוירופתיות פרוגרסיביות במבוגרים.
המסלול העיקרי בסינתזה של חומצות מרה מתחיל בשלב של הידרוקסילציה של כולסטרול בעמדה 7 על ידי פעולת האנזים CYP7A1 הממוקם ברטיקולום האנדופלזמי. מסלול זה בו CYP7A1 הוא השלב הראשון מכונה "המסלול הקלאסי או הנויטראלי", ויש מסלול חלופי בו השלב הראשון כרוך בהידרוקסילציה של כולסטרול בעמדה 27, על ידי האנזים המיטוכונדריאלי sterol 27-hydroxylase או CYP27A1. מסלול חלופי זה מכונה "המסלול החומצי", כאשר במכרסמים מסלול אחרון זה אחראי לכ-25% מכלל סינתזת חומצות המרה, באדם המסלול החומצי מהווה רק 6%. תוצרי הביניים של חומצות מרה הנוצרים על ידי פעילות CYP27A1 עוברים הידרוקסילציה בעמדה 7 על ידי האנזים oxysterol 7α hydroxylase או CYP7B1. למרות שהמסלול החומצי אינו מהווה מסלול עיקרי בסינתזת חומצת מרה באדם, נראה שיש לו חשיבות מעצם העובדה שמוטציה בגן CYP7B1 שהיא ממצא נדיר ביותר, גרמה בתינוק הפגוע לכולסטאזיס קשה עם צמקת הכבד.
שייר ההידרוקסיל בעמדה 3 נמצא באוריינטציה β וחייב לעבור אפימריזציה לאוריינטציה α, במהלך הסינתזה של חומצות מרה. האפימריזציה מתחילה בהתמרה של ה-3β-hydroxyl לקבוצת 3-oxo על ידי פעילות האנזים 3β-hydroxy-Δ5-C27-steroid oxidoreductase או HSD3B7. מוטציה בגן זה גורמת בילדים הלוקים בה למחלת כבד פרוגרסיבית המאופיינת על ידי צהבת כולסטאטית.
יצירתן של שתי חומצות המרה הראשוניות, cholic acid ו-chenodeoxycholic acid, נקבעת גם על ידי פעילות האנזים sterol 12α-hydroxylase או CYP8B1. תוצרי הביניים של פעולת האנזים HSD3B7 עליהם מגיב CYP8B1 הופכים ל-cholic acid, ואילו תוצרי הביניים עליהם אינו פועל HSD3B7, הופכים ל-chenodeoxycholic acid. לכן, פעילות הגן CYP8B1 תקבע גם את היחס ביןcholic acid ל-chenodeoxycholic acid.
וויסות ההומאוסטאזיס של חומצות מרה
חומצות מרה הן כאמור חומרים פעילי שטח (surfactants) בעלי אופי דטרגנטי, וכך הן יכולות להיות טוקסיות לתאים, מה שמחייב פיקוח מוקפד על ריכוזן.
חומצות מרה, ובעיקר שתי החומצות הראשוניות, cholic acid ו-chenodeoxycholic acid, מסוגלות לווסת את ביטוי הגנים המעורבים בסינתזה שלהן, בלולאה של משוב לאחור. הפענוח של מסלול מווסת זה התאפשר לאחר הבידוד של משפחת קולטנים הקרויים farnesoid X receptors, או FXRs. קולטנים אלה שייכים למשפחת-על של קולטנים הממוקמים על גרעין התא, הכוללת את משפחת הקולטנים של סטרואידים ושל הורמוני בלוטת התריס, כמו גם ה-liver X receptors או (LXRs), וכן את peroxisome proliferator-activated receptors או PPARs. שפעול FXR בכבד מעכב את הסינתזה של חומצות מרה. השפעול של FXR על ידי חומצות מרה במהלך ספיגתן במעי, מגביר את השעתוק והסינתזה של FGF19, שיעכב את יצירת חומצות המרה בכבד.
ישנם 2 גנים שזוהו כ-FXRα ו-FXRβ המקודדים ל-FXRs. באדם זוהו לפחות ארבעה איזופורמים של FXR, הנובעים מהגן FXRα, כתוצאה משפעול על ידי promoters שונים והשימוש בתהליכי splicing חלופיים. איזופורמים אלה FXRα3, FXRα2, FXRα1 ו-FXRα4. הגנים FXR מתבטאים ברמות הגבוהות ביותר במעיים ובכבד.
כמו כל הקולטנים במשפחת-על זו, הליגנד נקשר לקולטן בציטופלזמה ואז נודד הקומפלקס ליגנד-קולטן לגרעין התא, ויוצר הטרודימר עם חברים אחרים של המשפחה. FXR יוצר הטרודימר עם חברים של משפחת RXR. לאחר יצירת ההטרודימר, הקומפלקס נקשר לרצפים ספציפיים בגני מטרה הקרואים FXR response elements או FXREs, מה שגורם לביטוי מבוקר של גנים אלה. מטרה מרכזית אחת של FXR הוא השותף ההטרודימרי הגֶני הקטן SHP. שפעול של ביטוי SHP על ידי FXR מביא לעיכוב שעתוק הגנים של SHP. משמעותית לסינתזה של חומצות מרה, היא העובדה ש-SAP מדכא את ביטוי הגן CYP7A1 המקודד לאנזים cholesterol 7α-hydroxylase. אנזים זה הוא הקובע את קצב הסינתזה של חומצות מרה מכולסטרול במסלול הקלאסי.
ברפואה ההודית המסורתית, נהוג להפיק את התכשיר guggul מגזע העץ Commphfora mukul, לצורך הפחתת רמת כולסטרול. יכולת תכשיר זה להפחית כולסטרול נובעת מכך שמרכיב ליפידי בתמצית האמורה המכונה guggulsterone, מהווה אנטגוניסט של FXR. אך בנוסף להשפעתו של FXR, ה-guggulsterone ידוע כמשפעל של PXR או pregnane X receptor, חבר נוסף במשפחת העל של קולטנים בגרעין. PXR הוא קולטן מוכר של lithocholic acid ושל קודמנים (precursors) של חומצות מרה נוספות.
השפעול של PXR מוביל לדיכוי הסינתזה של חומצות מרה כתוצאה מהקשר המיידי עם גורם השעתוק הידוע כ-hepatocyte nuclear factor 4α או HNF-4α, מה שמונע מגורם השעתוק האחרון להגיב עם פקטור שעתוק נוסף הידוע כ-PGC-1α או PPARγ co-activator 1α, ובדיעבד ימנע את שפעול הגן CYP7A1.
הביטוי של גנים נוספים הקשורים לסינתזה של חומצות מרה, מבוקר על ידי פעילות FXR, שיכולה או לעודד גנים אלה או לדכא אותם. הגנים שפעילותם מדוכאת על ידי FXR בנוסף ל-CYP7A1 הם CYP8B1 המקודד לאנזים sterol 12α-hydroxylase, וכן SREBP-1c המקודד לאנזים הכרוך במטבוליזם של ליפידים, והגן SLC10A1 המקודד ל- solute carrier family 10, member 1. הגן האחרון מזוהה כ- Na+-taurocholate co-transporting polypeptide או NTCP, הקשור לקליטה של חומצות מרה בכבד, דרך הממברנה הסינוסואידאלית/בזולטראלית. אם כך, הדיכוי של ביטוי הגן NTCP על ידי חומצות מרה, מקטין את קליטת חומצות אלה ויגן על הכבד מהשפעה טוקסית של הצטברות עודף חומצות מרה.
בכבד, פעולת FXR מווסתת גם את ביטוי הגנים הכרוכים במטבוליזם של ליפופרוטאינים (דוגמת apoC-II), גנים הקשורים למטבוליזם של גלוקוזה (דוגמת PEPCK), וגנים הידועים בשמירה על תפקוד הכבד (דוגמת CYP3A4, שזוהה במקור כ-nifedipine oxidase: כידוע nifedipine הוא חבר במפחת התרופות החוסמות תעלות סידן).
חומצות מרה מדכאות את השעתוק של טרנספורטר נוסף שלהן המבוטא בממברנה הסינוסואידאלית/ בזולטראלית בכבד. טרנספורטר זה אינו תלוי בריכוזי +Na וידוע בכינוי organic anion transporting polypeptide 1B1 או OATP1B1, המקודד על ידי הגן SLCO1B1.
גנים, בנוסף ל-SHP, שדווקא מושרים על ידי FXR כוללים את BSEP או bile salt export pump, את MDR3 או multidrug resistance protein 3, וכן את הגן MRP2 או multidrug resistance associated protein 2. שני הגנים האחרונים קשורים לייצוא של תרכובות אורגניות או תרופות מהתאים. התפקיד הרגיל של MDR3 שהוא אחד המרכיבים של ה-ATP-binding cassette (ABC) family of transporters, הוא בטרנסלוקציה של פוספוליפידים דרך הממברנה הקנליקולארית של הפאטוציטים. לכן, משערים שהביטוי המוגבר של MDR3 המושרה על ידי חומצות מרה, נחוץ לאפשר להפאטוציטים להגיב לטוקסיות של חומצות המרה על ידי יצירה של מיצלה המורכבת מחומצות מרה, פוספוליפידים וכולסטרול. גם BSEP קשור התהליך ייצוא חומצות מרה מתוך הפאטוציטים והפחתת הנזק הטוקסי שעלול להיגרם לתאים אלה.
משמעות קלינית רבה ניתן למצוא בעובדה שרבים מהגנים עליהם משפיע FXR, משחקים תפקיד במספר מפגעים תורשתיים בכבד של כולסטאזיס. מוטציות ב-BSEP וב-MDR3, כרוכות במפגעים הידועים כ- familial intrahepatic cholestasis type 2 ו-familial intrahepatic cholestasis type 3, בהתאמה. מוטציות ב-MRP2 כרוכות בתסמונת Dubin-Johnson, שהיא צורה מורשת של היפר-בילירובינמיה.
ניתן בהחלט להתייחס לחומצות מרה כאל הורמונים סטרואידים, שכן הן מופרשות מהכבד ויש להם השפעה מטבולית ישירה ברחבי הגוף דרך הקולטנים הגרעיניים FXR וקולטנים על פני ממברנות תאים-TGR5.
צירקולציה אנטרו-הפאטית
כמויות גדולות של חומצות מרה מופרשות למעיים מדי יום, אך רק כמות קטנה יחסית אובדת מהגוף. זאת כיוון שבערך 95% מחומצות המרה המגיעות לתריסריון נספגות מחדש לדם בתוך המעי העקום (ileum). דם ורידי מה-ileum מגיע ישירות לווריד הפורטאלי, ומכאן דרך הסינוסואידים של הכבד, הפאטוציטים קולטים את חומצות המרה ביעילות רבה מתוך הדם הסינוסואידי, כאשר רק מעט מהן מתחמק מהכבד הבריא ועובר לצירקולציה הסיסטמית. החלק העיקרי של חומצות המרה עובר טרנספורט דרך ההפאטוציטים ומופרש לתוך ה-canaliculi. התוצאה הישירה של רה-צירקולציה אנטרו-הפאטית זו, היא שכל מולקולה של חומצת מרה מנוצלת מחדש בערך 20 פעם, לעתים אף פעמיים או שלוש פעמים במהלך של פאזת עיכול אחת.
מאגר חומצות המרה עובר 5-10 מחזורים של צירקולציה אנטרו-הפאטית מדי יום, כאשר הכבד בבריאות מלאה מפנה כל יום 20 גרם של מלחי מרה מהדם. פחות מ-1% מכלל המאגר של חומצות מרה בגוף נמצא בכל זמן נתון בדם ההיקפי, זאת כתוצאה מהיעילות הרבה של הטרנספורט הפורטאלי-כבדי של חומצות מרה. אלא שכאשר משתבשת מערכת הצירקולציה האנטרו-הפאטית רמת חומצות המרה בדם עולה משמעותית.
יש להדגיש שמחלת כבד עלולה לשנות באופן דרמטי את המתווה המתואר הזה של רה-צירקולציה. לדוגמה, תאי כבד חולים הם בעלי יכולת מופחתת למצות את חומצות המרה מהדם הפורטאלי כך שהנזק שיכול להיגרם בשל כל למערכת הקניקולארית יכול לגרום לדליפה של חומצות מרה לתוך הצירקולציה הסיסטמית. לכן חיוני למדוד רמת חומצות מרה בדם שהוא מדד קליני רגיש למחלת כבד.
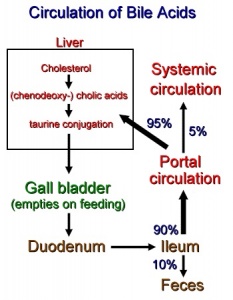
המסלול האֶנטֶרוֹ-הֶפּאטי התקין של חומצות מרה
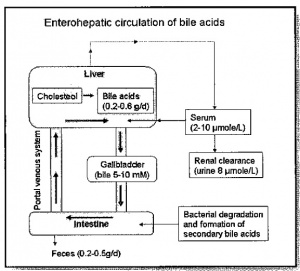
כמויות של חומצות מרה העוברות במסלול אנטרו-הפאטי תקין
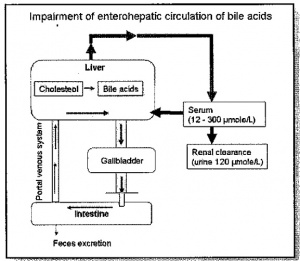
כמויות של חומצות מרה העוברות במסלול אנטרו-הפאטי פגום



המתווה והפיקוח של הפרשת המרה
הפרשת המרה נמוכה ביותר בעת צום, כאשר רוב נוזל המרה מוסט לכיס המרה לצורך ריכוזו. כאשר בליל המזון המעוכל בחלקו בצאתו מהקיבה לתריסריון הידוע כ-chyme נכנס למעי הדק, חומצה ושומני מזון וחלבונים המעוכלים באופן חלקי, מעודדים הפרשה של 2 הורמונים, cholecystokinin או CCK ו-secretin. הורמוני מעי אלה משפיעים באופן משמעותי על הפרשת הורמוני הלבלב, ובנוסף יש להם תפקיד בהפרשת ושפיעת המרה לתוך המעי: הגירוי המשמעותי ביותר להפרשת cholecystokinin היא נוכחות שומנים בתריסריון, ואילו הגירוי המשפיע על הפרשת secretin היא דרגת החומציות בתריסריון. השפעת secretin על מערכת המרה היא בגירוי של תאים בצינורות המרה להפריש ביקרבונאט ומים, מה שמרחיב את נפח המרה ומגדיל את קצב שפיעתה אל המעי.
שתי חומצות המרה השכיחות ביותר בנוזל המרה הן chenodeoxycholic acid (המהווה 45%) ו-cholic acid (המהווה 31%). לפני ש-2 חומצות מרה ראשוניות אלו מופרשות לתוך הלומן הקניקולארי, הן עוברות קוניוגציה דרך הקשר האמידי בקצה הקרבוקסי-טרמינאלי שלהן עם גליצין או עם טאורין. קוניוגציה זו מגבירה את האופי האמפיפאתי שלהן, מה שמאפשר להן להיות מופרשות ביתר קלות ואף להיות פחות טוקסיות. קוניוגאטים של חומצות מרה הם המרכיבים העיקריים של נוזל המרה.
לאחר ההפרשה על ידי הכבד, חומצות המרה נכנסות לנימי המרה (קנליקולי) המתחברים לצינוריות המרה (bile ductules) ליצירת צינורות המרה (bile ducts). חומצות המרה נישאות מן הכבד בצינורות אלה בדרכן לכיס המרה, בו הן נאגרות לצריכה עתידית. בכיס המרה חלל ריכוז משמעותי של חומצות המרה עד פי-1,000.
בתוך המעיים מגיבים חיידקי מעי עם חומצות המרה הראשוניות, ואלו עוברות תהליך של דה-קוניוגציה המרחיק את שיירי הגליצין והטאורין. אחוז קטן של חומצות המרה החופשיות יכול להיות מופרש בצואה או להיספג מחדש על ידי המעי ולחזור לדם בדרכו חזרה לכבד. חיידקים אנארוביים המצויים במעי הגס, יכולים להסב חומצות מרה ראשוניות, ולהפכן לחומצות מרה שניוניות, כמו deoxycholate שמקורו ב-cholate או lithocholate שמקורו ב- chenodeoxycholate. חומצות מרה ראשוניות כמו גם שניוניות נספגות מחדש במעי וחוזרות לכבד דרך הצירקולציה הפורטאלית. אכן, עד 95% מכלל חומצות המרה המסונתזות בכבד נספגות דרך ה-ileum הדיסטאלי וחוזרות לכבד.
תפקיד רגולטורי לחומצות מרה במטבוליזם
חומצות מרה זוהו כמעורבות ב-4 תהליכים פיזיולוגיים ראשוניים משמעותיים:
- הסינתזה שלהן והפרשתן בצואה מייצגים את המנגנון המשמעותי הבלעדי של הרחקת עודף כולסטרול מהגוף.
- חומצות מרה ופוספוליפידים ממיסים כולסטרול בנוזל המרה, ועל ידי כך מונעים את שקיעת כולסטרול בכיס המרה.
- חומצות מרה מסייעות לעיכול של טריגליצרידים על ידי יצירת תחליב המאפשר לאנזים lipase ממקור הלבלב לפרק שומן דיאטתי זה.
- חומצות שומן מסייעות לספיגה במעי של ויטמינים מסיסי-שומן (A, D, E, ו-K).
בנוסף לתפקידים של חומצות מרה בתחלוב שומנים במעי ושפעול של FXR, הן גם משתתפות במספר תהליכים מושרים על ידי איתותים על ידי שפעול של JNK או c-JUN N-terminal kinase, כמו גם שפעול של מסלול MAMK או mitogen-activated protein kinase. חברים אחרים במשפחת קולטני הגרעין המשופעלים על ידי חומצות מרה בנוסף ל-PXR שכבר הוזכר למעלה, הם CAR או constitutive androstane receptor, וכן VDR או vitamin D receptor. כמו כן קולטן נוסף המשופעל על ידי חומצות מרה שעשוי להשפיע על בקרת ההשמנה, הוא TGR5 או transmembrane G-protein coupled bile acid receptor 1, הידוע גם כ-GPR131. שפעול של הקולטן האחרון ברקמת השומן החומה מביא לשפעול של UCP1 או uncoupling protein 1 (ידוע גם כ-thermogenin), המביא לצריכת אנרגיה מוגברת.
שינויים ברמת חומצות מרה בתרחישים קליניים שונים
רמת חומצות מרה מוגברת במחלות כבד בהן יש שינויים מבניים בכבד כתוצאה מאי היכולת של הכבד הפגוע לקלוט ביעילות חומצות מרה מהדם הפורטאלי. מדובר במפגעי כבד כמו הפאטיטיס נגיפי, מחלת כבד אלכוהולית, anicteric liver disease, צמקת הכבד, כולסטאזיס, קרצינומה הפאטו-צלולרית, הפאטומה ראשונית, גרד, בחילה והקאות בהריון, protracted diarrhea of infancy, תסמונת Reye, פגיעה כבדית על רקע צריכת תרופות או חשיפה לכימיקאלים שונים, neonatal hepatitis syndrome, לייפת כיסתית (cystic fibrosis) וחסימה מרתית (biliary atresia). לעומת זאת, מחלות כבד מטבוליות כמחלת Gilbert, תסמונת Crigler-Najjar או תסמונת Dubin-Johnson, אינם גורמים לעלייה ברמת חומצות מרה.
מפגעים מטבוליים מוּלדים הקשורים לביוסינתזה ולמטבוליזם של חומצות מרה, מסווגים כמפגעים ראשוניים או שניוניים. מפגעים ראשוניים נובעים מחסר באנזימים האחראיים לקטליזה של סינתזה של חומצה כולית וחומצה כנודאוקסיכולית. מפגעים משניים של חומצות מרה מוגדרים כפגמים מטבוליים המשפיעים על סינתזה של חומצות מרה שאינם נגרמים מפגיעה באנזימים האחראיים לסינתזה זו. פגמים משניים של מטבוליזם של חומצות מרה כוללים מפגעים פראוקסיזומיים כגון תסמונת Zellweger, וכן תסמונת Smith-Lemli-Opitz הנובעת מחסר של האנזים dehydrocholesterol reductase-7 או DHCR7.
| אנזים פגום | סמל הגן | המפגעים הקליניים |
|---|---|---|
| cholesterol 7α-hydroxylase | CYP7A1 | אין בעיה בתפקודי הכבד, אך רמת סך-כולסטרול וכולסטרול-LDL גבוהה ביותר. יש הופעת אבני מרה בגיל צעיר, כמו גם מחלת עורקים היקפיים. סטאטינים אינם משפיעים על רמות כולסטרול. |
| sterol 27-hydroxylase | CYP27A1 | אי-תפקוד נוירולוגי פרוגרסיבי. הופעת כולסטאזיס נאונטאלי, ירוד דו-צדדי, שלשולים כרוניים. |
| oxysterol 7α-hydroxylase | CYP7B1 | מקרה אחד דווח בשנת 1998, של תינוק בן-10 שבועות עם כולסטאזיס פרוגרסיבי, טחול וכבד מוגדלים, צמץ כבד, וכשל כבדי, עם רמות ALT ו-AST מוגברות ביותר. לאחרונה נמצא שבמספר פרטים הסובלים משיתוק רגליים ספסטי מורש (SPG) אוטוזומאלית-רצסיבית מטיפוס 5A, התגלו מוטציות בגן CYP7B1, אם כי מספר מקרים אלה מהווים רק כ-1% מכלל מקרי SPG. |
| 3β-hydroxy-Δ5-C27-steroid oxidoreductase | HSD3B7 | המפגע השכיח ביותר בסינתזה של חומצות מרה, המתבטא בתסמינים קליניים הטרוגניים, הכוללים צהבת פרוגרסיבית, כבד מוגדל, גרד, בעיות ספיגה במעי עם שלשולים שומנים (steatorrhea), חסר בוויטמינים מסיסי שומן ורככת עצמות. |
| Δ4-3-oxosteroid 5β-reductase | AKR1C4 | תסמינים קליניים דומים לאלה שבחסר HSD3B7, אם כי התסמינים מופיעים מוקדם יותר. הילדים הפגועים יסבלו ממחלת כבד חמורה יותר עם החמרה מהירה לצמקת ולמוות אם הטיפול אינן מיידי ונחוש. בין התסמינים עלייה ניכרת ברמת AST ו-ALT, עלייה ברמת בילירובין מצומד, והפרעות קרישה. |
| 2-methylacyl-CoA racemase | AMACR | לראשונה תואר בשלושה מבוגרים עם נוירופתיה סנסורית מוטורית, וכן אובחן מפגע זה במקרה של תינוק בן 10 שבועות עם חסר ניכר בוויטמינים מסיסי-שומן, hematochezia (צואה בגוון דמי) ומחלת עימדון כבד מתונה. |
עימדון מרה (כולסטזיס) תוך-כבדי בהריון (ICP או Intrahepatic cholestasis of pregnancy)
מפגע המתבטא בגרד עז שמתחיל בשבוע 25-32 להיריון, או אף קודם לכן. עלייה ברמת חומצות המרה בדם עם ממצא היסטולוגי בביופסית כבד המתאים לעימדון מרה (כולסטזיס). התסמינים נעלמים לאחר הלידה אך עלולים להופיע בהריונות הבאים. הגרד חמור ביותר בכפות הידיים והרגליים ומחמיר בלילה. צהבת תופיע ב-10-25% מהמקרים כשבועיים עד 4 שבועות אחרי הופעת הגרד. עימדון המרה הקשה עלול לגרום לצואה שומנית (סטאטוראה) עם ירידה בספיגת ויטמיני ADEK. רמת חומצות מרה ב-ICP היא לפחות פי-3 גבוהה יותר מהרמה התקינה, אם כי רמה זו עלולה להיות פי-10 עד פי-100 גבוהה יותר.

מפגע זה פוגע יותר בנשים יוצאות סקנדינביה או צ'ילי, אך לא נמצא קשר לקבוצות HLA. אבחנה - עליית רמות חומצות מרה בסרום מוכיחה את האבחנה. מפגע זה פוגע יותר בנשים יוצאות סקנדינביה או צ'ילי, אך לא נמצא קשר לקבוצות HLA. אבחנה - עליית רמות חומצות מרה בסרום מוכיחה את האבחנה. מחקר פיני שכלל מעל 10,000 נשים מצא שמטופלות הסובלות מ-ICP נמצאות בסיכון מוגבר להתפתחות אבני מרה, דלקת לבלב חריפה, דלקת כבד נגיפית מסוג C ושחמת שלא על רקע אלכוהולי.
חומצות מרה וסרטן המעי הגס
נראה שחומצות מרה הן בעלות משמעות בהתרחשות סרטן המעי הגס: חומצת המרה deoxycholic acid מוגברת בתכולת המעי הגס בתגובה לדיאטה עתירת שומנים. באוכלוסיות עם שיעור גבוה של סרטן המעי הגס, מוגבר ריכוז חומצות השומן בצואה, מה שמרמז לחשיפה מוגברת של הלוּמֶן של המעי הגס לריכוזים מוגברים של חומצות המרה עלול לשחק תפקיד בהתפתחות סרטן המעי הגס.
בהשוואה ייחודית, הריכוז של deoxycholic acid בצואה של תושבי דרום אפריקה הילידים שהדיאטה שלהם דלה בשומנים, היא 7.3 ננומול' לגר' משקל רטוב של צואה. לעומת זאת, אפרו-אמריקנים בארה"ב שהדיאטה שלהם עתירת שומנים, מכילים בצואה חומצת מרה זו בריכוז של 37.5 ננומול' לגר' משקל רטוב של צואה. ואמנם, ילידים דרום אפריקנים לוקים בשיעור נמוך ביותר של סרטן המעי הגס, העומד על מקרה אחד ל-100,000, בה בשעה ששיעור סוג זה של סרטן בקרב אפרו-אמריקנים עומד על 72 מקרים לאוכלוסיה של 100,000.
מחקרים אפידמיולוגיים וניסויים מצביעים גם כן על תפקיד חומצות שומן בהתרחשות סרטן המעי הגס. סקירה של Bernstein וחב' שהקיפה 12 מחקרים מצביעה שחשיפת תאי מעי גס לרמות גבוהות של deoxycholic acid מגבירה יצירת תוצרים פעילי חמצון המגדילים את העקה החמצונית, ו-14 מחקרים שפורסמו מראים שחשיפת תאים לחומצות מרה מגבירים את הנזק ל-DNA התאי. תאים שורדים בהם לא חל תיקון של הנזק ב-DNA, עלולים בחלוקתם להביא ליצירת תאי-בת עם מוטציות מסרטנות.
מחקר ניסויי אחד, בו הוזנו עכברים משך 8-10 חודשים עם דיאטה מועשרת בחומצה דאוקסיכולית אקסוגנית, מצא שבין 18 עכברים כאלה, ב-17 התפתחו גידולים במעי הגס, מתוכם ב-10 הגידולים היו ממאירים. לעומתם לא נמצאו כל גידולים בעכברי ביקורת שניזונו מדיאטה רגילה בה כמות החומצה הדאוקסיכולית הייתה רק 10% בהשוואה לקבוצת המדגם.
תרופות המשפיעות על רמת חומצות מרה
תרופות שמעלות את רמת חומצות המרה בנסיוב: ציקלוספורין, isoniazide, מתוטרקסאט, rifampin. תרופה המפחיתה רמת חומצות מרה בנסיוב: cholestyramine.
תרופות המעלות רמת חומצות מרה בצואה: cholestyramine, נאומיצין. תרופות המפחיתות רמת חומצות מרה בצואה: clofibrate, rifampin וכן דיאטה עשירה בפחמימות.
שיטות למדידת רמת חומצות מרה
בעבר היה נהוג למדוד רמת חומצות מרה בשיטות אנזימטיות, אך בשנים האחרונות גוברת הנטייה למדידה ישירה של חומצות מרה בשיטות של RIA או GLCאו הפרדה וכימות בעזרת HPLC. השיטות האנזימטיות מודדות את סך חומצות המרה החופשיות ואלה שעברו קוניוגציה, פרט לחומצות מרה עם שייר סולפאט בעמדה C3.
בין שיטות המעבדה לקביעת רמת סך חומצות מרה (TBA או total bile acid), מקובלות במעבדות שיגרה שאינן מצוידות במכשור של GLC או HPLC לפעילות יומיומית, שיטות המדידה האנזימטיות:
- שיטה אנזימטית (דור שלישי)-כאן משתמשים באנזים 3α-HSD או -3α-hydroxysteroid dehydrogenase, כדי לקטלז את ריאקציה החמצון הממירה את שייר הידרוקסיל בעמדה 3α של כל חומצות המרה, לשייר 3-keto עם יצירה NADH מ-+NAD. תוצר הביניים NADH מגיב עם nitrotetrazolium blue או NBT, ליצירת הצבע formazan על ידי האנזים diaphorase. עוצמת הצבע הנוצר נמדדת באורך גל 540 ננומטר, והיא פרופורציונית לריכוז מלחי המרה בדגימת הנסיוב.
- שיטת המִחזוּר האנזימטי (enzyme cycling method, דור חמישי)-שיטה זו מאפשרת signal amplification על ידי מִחזור פעילות האנזים. בשיטה זו, מולקולות חומצות המרה בנסיוב, עוברות מחזורי חמצון וחיזור נשנים וחוזרים, על ידי האנזים -3α-HSD כמו בשיטה הקודמת, עם הצטברות בו-זמנית של הקו-אנזים המחוזר thio-NADH שניתן לגלות באורך גל 405 ננומטר.

כפי שניתן לראות בתרשים בריאקציה המקדימה אנזים זה מקטלז את ריאקצית החמצון בנוכחות הקו-אנזים +thio-NAD, ליצירת חומצות מרה מחומצנות, וצורת קו-אנזים מחוזרת-thio-NADH. מצד שני, בריאקציה ההפוכה, האנזים מקטלז את ריאקצית החיזור בנוכחות של עודף קו-אנזים NADH להפיכת חומצות מרה מחומצנות בחזרה לחומצות מרה, המוכנות כעת למחזור השני של ריאקצית חמצון. בשיטה זו מקבלים רגישות גבוהה בהרבה של גילוי חומצות מרה, כאשר שיעור יצירת thio-NADH עומד ביחס ישר לריכוז חומצות המרה בדגימת הנסיוב.
היתרון של השיטה האחרונה על השיטה האנזימטית הקודמת, הוא במספר פרמטרים: ניתן לדגום ישירות מהנסיוב ללא צורך בליופיליזציה של הדגימה לקבלת אבקה כחומר מוצא; ניתן להשתמש בנפח דגימה מזערי של 3-5 מיקרוליטר בהשוואה ל-20 מיקרוליטר בשיטה הקודמת; אין הפרעה לריאקציה האנזימטית מדגימה ליפמית או המוליטית, שעה שהשיטה האנזימטית הקודמת כן מושפעת משני פרמטרים אלה.
הוראות לביצוע הבדיקה
יש לדגום דם לאחר צום של 12 שעות במבחנה כימית (פקק אדום או צהוב). ניתן לשמור את הנסיוב יום בטמפרטורת החדר, עד שלושה ימים מקורר או עד שבוע מוקפא. יש להקפיד שנסיוב לא יהיה מזוהם על ידי חיידקים. אין להשתמש במבחנות עם נוגדי קרישה, שכן יש העדפה לביצוע המדידה בנסיוב ולא בפלזמה. אין להקפיא ולהפשיר את דגימת הנסיוב יותר מפעמיים. צמיחת חיידקים כלשהי בדגימת הנסיוב צפויה לשנות את ריכוז חומצות המרה. ביצוע מדידה של רמת חומצות מרה בפלזמה, יפיק תוצאות הנמוכות בכ-10% בהשוואה למדידה שלהן בנסיוב.